Icons of Evolution: Why are there so many Darwin’s finches?
Uncover the remarkable new development in the understanding of the evolution of Darwin's finches, discovering evidence of extinctions for the first time.
Published on 23rd October 2024
Darwin’s finches are an iconic example of adaptive radiation, where a single common ancestor rapidly diversified into 18 species over just 1–2 million years. Their evolution occurred dramatically, with variation in their size, beak shape, and behaviour being the most extreme. What’s more, variation mostly occurred without genetic divergence resulting in unviability between lineages, something that is common in other radiations.
Renowned evolutionary biologists Peter and Rosemary Grant used their long-term field research on Daphne Major, a large island within the Galápagos archipelago, and recent genomic analyses to challenge the traditional model of adaptive radiation. The full paper, published as part of a Special Issue on 'A Global Perspective on Adaptive Radiation' in the Evolutionary Journal of the Linnean Society, can be found on the journal's homepage.
It is for several reasons that Darwin’s finches make good models for understanding adaptive radiation. They’re easy to observe and capture, they mature quickly, their habitat is mostly intact, and species can be found both together and separately. More importantly, human activity has not led to the extinction of a single species. In short, species-level radiation (where species evolve from one into many) is pristine.
However, despite being a textbook example of adaptive radiation, questions remain: why did species diversify so rapidly, and how did they do it?
The Ecology of Adaptive Radiation
Traditionally, there has been widespread agreement over the allopatry-then-sympatry (ATS) model of adaptive radiation, whereby genetic differentiation of geographically isolated populations (allopatry) is followed by further differentiation in coexisting populations (sympatry). Natural selection during the isolation phase reduces competition with other species, and thus the risk of interbreeding, leading to speciation.
Evidence for this theory comes in the form of the medium ground finch (Geospiza fortis) and the heritability of beak size. A drought on Daphne Major in 1977 led to a decline in its population due to size-selective mortality; their beaks were too small to open the large and hard fruits that were available. As a result, beak size in the next generation increased, supporting the allopatry half of the theory. Later, a population of the large ground finch (Geospiza magnirostris), a competitor species, established itself on Daphne Major. Eventually, it out-competed G. fortis after a second drought on the island. Morphological differences arose between the two species through competitively induced character displacement, supporting the sympatry half of the model.

Medium ground finch (Geospiza fortis) © Peter and Rosemary Grant

Large ground finch (Geospiza magnirostris) © Peter and Rosemary Grant
But why 18 species and not just two? And what sets the limit for number of species? For multiple speciation events to occur, environments offering diverse ecological opportunities (multiple niches) are required, as well as sufficient time and genetic potential. Given the large number of islands in the Galápagos archipelago, and the proximity of the islands to one another, the potential for evolution is high.
Islands that don’t have these conditions do not have multiple species: the isolated Cocos Island to the north of the Galápagos has only one. Given this ecological potential, the number of species that evolve is limited by a combination of factors, including resource availability, competition and spatial isolation.
Reproduction and Hybridisation
For species to coexist long term they must be reproductively and ecologically ‘isolated’. Among Darwin’s finches, plumage and courtship behaviour play a lesser role in species isolation than in other passerine birds. Instead, morphology and song are stronger means of distinguishing species in a shared habitat. Using stuffed museum specimens, ground finches were shown to recognise their species through appearance alone, whilst song playback showed this was also possible through acoustic signals. This evidence, paired with the understanding that bird song is taught to nestlings by their fathers, shows that reproductive barriers within this genus are both genetically and culturally inherited.
In some species of Darwin’s finches, these barriers to interbreeding are crossed. Introgressive hybridisation or introgression, where the genes of one taxon are incorporated into the gene pool of another, is a phenomenon seen in both the ground and tree finches in the Galápagos archipelago since 1976. Though not typically found in birds, the discovery of introgression in Darwin’s finches questions the fitness of these hybrid species. When hybrid individuals breed with their parental species, no costs to fitness were found. As such, speciation is driven by natural selection, with reproductive isolation being a by-product of morphological changes.
Yet, in recent years hybrid speciation has occurred in Galápagos finches, something that is theoretically unexpected. A male cactus finch (G. scandens) immigrated to Daphne Major in 1981 and bred with a female medium ground finch in 1983. Backcrossing initially occurred, resulting in introgression, but by the third generation there was no outbreeding, and this new lineage was acting as an incipient or 'almost' species, without reduced fitness.

Cactus finch (Geospiza scandens) © Peter and Rosemary Grant
What’s more, the merging of G. fortis and G. scandens on Daphne Major is on a speciation trajectory; individuals in 2012 were 1.5x more similar than they were in the 1980s, both genetically and phenotypically. As such, the classical ATS theory of adaptive radiation is insufficient, failing to explain the source of genetic variation. Hybridisation increases genetic variance and creates new species, resulting in high heritability of new traits and increasing the potential for evolution. In Daphne Major’s finches this is seen in beak traits, resulting in rapid adaptive radiation.
Finches Travel Through Time
Alongside genetic factors, environmental change contributed to the radiation of finches in the Galápagos. The archipelago is not a stable ecosystem; volcanic activity and changing sea levels over glacial periods led to the formation and destruction of new islands. A net increase in island number created countless new niches, which in turn led to an increase in finch species. It appears that the invasion of these islands by South American plants and arthropods around 250,000 years ago further increased the number of ecological opportunities available, which may account for the evolution of seven species within 50,000 years. The cactus finch (G. scandens) evolved around this time, perhaps triggered by the arrival of the Opuntia prickly pear cactus after a period of rapid warming following the end of the Last Glacial Maximum (LGM). A dated phylogeny is needed to say this for sure, which you can read about in another of our Special Issue papers.
The Extinction Problem
Whilst the simplest explanation for radiation assumes all species that have existed still exist, this is not always the case. Analysis of fossil evidence can estimate ancestral phenotypes, yet sufficiently old fossils of Darwin’s finches do not exist. Therefore, indirect evidence must be used, assuming extinction left a mark on finch evolution.
Take the warbler finch (Certhidea), which split from the ancestral state (assumed to be the seed-eating dull coloured grassquit (Asemospiza obscura)) around 1.4 Mya. Though much smaller than Asemospiza, its beak is longer and pointier due to its insectivorous diet. As both beak traits changed in opposite directions, the evolution of the warbler finch likely occurred over a long period, making it likely that extinctions occurred during this time.
Since the initial split of the ancestral lineage, the next stage of radiation occurred a quarter to half a million years later. One branch led to the vegetarian finch (Platyspiza crassirostris), and the other to all other species in the radiation. Beak size and shape between the vegetarian and warbler finch span the full range of morphological differences, suggesting intermediate species must have existed.
Furthermore, several species that have evolved in the last 400–200,000 years possess beak traits in the mid-range of the radiation, indicating that the evolutionary trajectory has reversed on itself. Given they occupy the mid-point, several species must have existed in the past within this range.
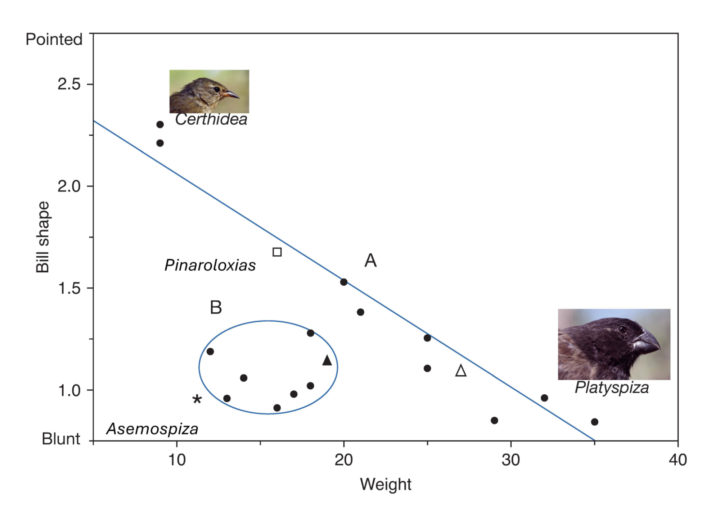
Mean morphology of all Darwin's finches, showing the variance in bill shape in relation to body size in most species (Group A). Shape becomes blunter as body size increases. Group B (circled in blue) shows a deviation from this relationship. © Peter and Rosemary Grant
Overall, this remarkable new evidence of extinction of species through geophysical factors and competition suggests the adaptive radiation occurred not just through natural selection, but also through the selective replacement of species over time. The slow beginning and substantial morphological changes in their evolution opposes the theory of rapid evolution after a founding species invades a new environment. Whilst early differentiation may have occurred, a lack of evidence for extinct species with these features makes it hard to know for sure.
Macrocosm to Microcosm
Though we cannot travel back in time, the present illuminates the past. When considering Daphne Major as a case study, long periods of selection associated with extreme events, such as prolonged drought, followed by long periods of little to no selection, act as an example of small-scale patterns on a largescale. That is, the stasis of evolution for a millennium followed by rapid evolution and speciation when confronted with new environments. The divergence of fortis on Daphne Major is the best example, whereby the fluctuations in average beak size across just 40 years is equivalent to the divergence of 8/11 populations of fortis on other islands in the archipelago.
In short, this evidence paints a picture of speciation that is rapid and more limited by ecological opportunities than genetic potential. Combining the effect of hybridisation in the generation of new species, Daphne Major appears to be acting as a microcosm to the Galápagos macrocosm. The Grants therefore propose that the traditional ATS theory should be replaced by a competition-selection-hybridisation model to account for the different processes involved in the generation of new species.
About the Paper Authors

© Peter and Rosemary Grant
Professor Peter Grant FLS FRS and Professor Rosemary Grant FRS, renowned evolutionary biologists, have studied Darwin’s finches on the Galápagos islands since 1973. Their fieldwork is designed to understand the causes of an adaptive radiation. Their work is a blend of ecology, behaviour and genetics, collaborating with investigators to estimate phylogenetic relations among the species of finches and their relatives on the continent and in the Caribbean.
Rosemary was initially trained at the University of Edinburgh, received a PhD degree from Uppsala University, and was a Research scholar and Professor in the Department of Ecology and Evolutionary Biology at Princeton University until she retired from teaching in 2008. Peter is the Class of 1877 Professor Emeritus in the same Department, having trained at Cambridge University and the University of British Columbia. Before joining Princeton in 1986, he taught at McGill University and the University of Michigan.

Blog Editor
Compiled and edited by Georgia Cowie, Journal Officer (Linnean Society). © The Linnean Society
